






.jpg)
.jpg)
.jpg)

















Эта новая формулировка определения направления (в терминах проекций на сетчатки относительно проекций носа, а не в терминах абсолютной ретинальной локализации) решает некоторые проблемы, которые совершенно непонятны в рамках традиционной теории. Проблема мгновенной адаптации к искажающим призмам перестает быть проблемой, как только мы признаем, что ретинальная проекция наблюдателя смещается призмами тем же самым образом, что и проекции объектов вне тела. Здесь есть абсолютное ретинальное смещение, но нет смещения относительно проекций носа и орбит. Проблема косоглазия также может быть успешно разрешена с помощью предложенной интерпретации. В случае косоглазия один глаз постоянно находится под определенным углом к другому (рис. 3.17.). Это означает, что фовеа двух глаз постоянно направлены на различные точки в пространстве. Несмотря на это, оба глаза видят одни и те же объекты и в одних и тех же направлениях — нечто совершенно невероятное, если только мы не предположим, что информация, определяющая восприятие дистального положения, заключается в относительном положении проекции объекта и проекции самого наблюдателя.
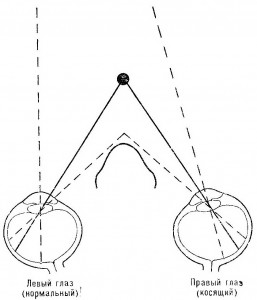
Рис. 3.17. Случай косоглазия. Фовеа глаз постоянно направлены в различных направлениях. Ретинальные проекции некоторого объекта падают на разные («некорреспондирующие») участки сетчаток обоих глаз. Это происходит также и с проекциями носа.
Предложенная здесь гипотеза может даже объяснить результаты двух излюбленных демонстраций сторонников теории эфферентной копии. Рассмотрим случай пассивного смещения глазного яблока. Во-первых, я вообще отрицаю, что кто-либо может создать пассивное движение глаза, которое будет похоже на активное движение глаза в отношении своих зрительных эффектов. Нормальное движение глаза представляет собой вращение, оно не вызывает изменения в относительном проекционном положении себя и объектов. Как вы сами можете убедиться, пассивные движения глаз приводят к изменениям относительного проекционного положения наблюдателя и других объектов, что, согласно предложенной нами гипотезе, является условием восприятия движения. Что касается результатов, полученных в ситуации ограничения подвижности глазных яблок, то я не могу сказать лучше Уильяма Джеймса:
«Мне очень хотелось, но сам я не смог успешно повторить этот опыт — не знаю, по какой причине. Но даже по отношению к тем случаям, когда он удавался, я должен заметить, что его условия слишком сложны, чтобы можно было с уверенностью делать те выводы, которые делает профессор Мах. Замазка и давление, которое она оказывает на глазное яблоко, должны приводить к возникновению достаточно сильных периферических ощущений. Эти ощущения могут объяснить любые проявления ошибочного восприятия положения глазных яблок совершенно независимо от чувства иннервации, существование которого допускает профессор Мах» (Джеймс, 1890) [7].
Психологи склонны думать об «Я» либо как о бессмысленной абстракции, либо как о сложной проблеме, требующей глубокого размышления. Сведение «Я» к зрительной проекции носа на сетчатку обычно вызывает радостное оживление и насмешки слушателей. Даже Гибсон (1950), который сделал для понимания роли проекции носа больше, чем какой-либо другой психолог, писал об этом со своего рода защитной уклончивостью. Я хотел бы сказать здесь несколько слов в защиту носа. Среди самых разных видов животных с камерными глазами нет ни одного, чьи глаза не получали бы проекцию носа или какой-нибудь другой часта тела. У всех представителей видов с развитым панорамным зрением панорама прерывается проекцией самого животного. У представителен хищных видов, отличающихся максимальной фронтальностью положения глаз и максимальным бинокулярным перекрытием, всегда есть проекция себя. Это не может быть просто биологической случайностью. Проекция частей тела на сетчатку должна иметь важное функциональное значение, иначе она бы не сохранилась у столь многих эволюционно различных форм. В самом деле, предложенная здесь гипотеза о восприятии положения имеет то достоинство, что она придает смысл факту, который иначе кажется длительной биологической ошибкой (Уолс, 1942) (см. рис. 3.18) [8].
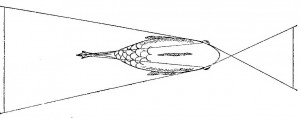
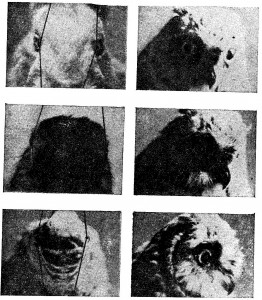
Рис. 3.18. Зрительное поле различных животных. У зайца (слева вверху и в середине) и вальдшнепа (слева внизу), которые являются жертвами для многих других животных, развилось исключительно широкое зрительное поле, охватывающее до 200°. Вследствие этого как заяц, так и вальдшнеп могут одновременно видеть свой нос (клюв) и хвост. Рыба имеет общее зрительное поле около 360°. Значительная его часть занята телом и плавниками рыбы. Сова имеет зрительный угол 110° с бинокулярным перекрытием, ее глаза направлены прямо вперед. Однако у этой птицы есть специальный назальный хохолок, который занимает центральные 30° ее зрительного поля.
ретинальные проекции – предыдущая | следующая – стимульные последствия движения
Психическое развитие младенца. Содержание.